Utover genomet - et ikke-reduksjonistisk perspektiv på utvikling
Jonathan M. 2.juni 2011.
Oversatt herfra. (kursivering og understreking ved oversetter)
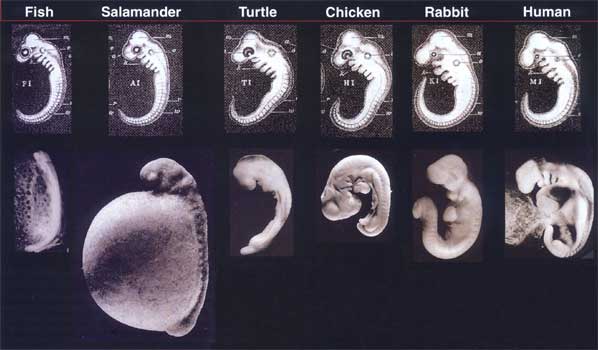 Et vanlig tema som ofte gjennomsyrer diskusjoner knyttet til embryologi og utviklingsbiologi - spesielt, innen (men ikke begrenset til) ID-kretser - er ideen om at en organismes DNA-sekvens ikke alene er ansvarlig for utviklingen av den tredimensjonale strukturen og arkitekturen som er funnet i celler, organer og kroppsplaner.
Et vanlig tema som ofte gjennomsyrer diskusjoner knyttet til embryologi og utviklingsbiologi - spesielt, innen (men ikke begrenset til) ID-kretser - er ideen om at en organismes DNA-sekvens ikke alene er ansvarlig for utviklingen av den tredimensjonale strukturen og arkitekturen som er funnet i celler, organer og kroppsplaner.
Som forklart her av biolog Jonathan Wells, inneholder hver celle i en utviklende organisme (med ett eller to bemerkelsesverdige unntak som T- og B-lymfocytter) - enten i det potensielle hodeområdet til et embryo eller i den potensielle haleregionen til et embryo - nøyaktig den samme DNA-sekvensen (et fenomen kjent som 'genomisk ekvivalens'). Differensielt genuttrykk refererer til aktivering av forskjellige gener i en organismes respektive steder på spesifiserte tidspunkter under embryogenesen.
Hodeceller må slå på forskjellige gener i forhold til haleceller, og de 'vet' hvilke gener de skal slå på fordi de mottar informasjon om deres romlige beliggenhet utenfor seg selv - og dermed utenfor DNA-sekvensens virke.
Av kritisk betydning for genuttrykksmønstre er DNA-bindende transkripsjonsfaktorer. Disse spesialiserte proteiner er ansvarlige for uttrykk eller undertrykkelse av gener ved å binde spesifikke nukleotidsekvenser som finnes i deres promotorer. Påfølgende interaksjon med deres anerkjente sekvenser resulterer i en kaskade av hendelser. Dette innebærer ofte endringer i kromatin-arkitektur, som fører til montering av et aktivt transkripsjonskompleks (Cosma et al. 1999). Det som er viktig å merke seg er at de typer transkripsjonsfaktorer som er til stede i en celle ikke i seg selv er tilstrekkelige til å definere dets respektive spekter av genuttrykk.
 DNA-metyleringsmønstre, i tillegg til noen få andre mekanismer, tjener som et elegant 'epigenetisk minne' -system for å sikre irreversibiliteten til differensial-genuttrykk (Bird, 2002). Faktisk bemerker Bird-artikkelen at:
DNA-metyleringsmønstre, i tillegg til noen få andre mekanismer, tjener som et elegant 'epigenetisk minne' -system for å sikre irreversibiliteten til differensial-genuttrykk (Bird, 2002). Faktisk bemerker Bird-artikkelen at:
En modell 'kun av transkripsjonsfaktorer' ville forutsi at genuttrykksmønsteret til en differensiert kjerne ville være fullstendig reversibel ved eksponering for et nytt spekter av faktorer. Selv om mange aspekter av uttrykk kan omprogrammeres på denne måten (Gurdon 1999), er noen merker av differensiering tydeligvis så stabile at nedsenking i en fremmed cytoplasma ikke kan slette minnet.
Rinn et al. (2006) bemerker at:
Fibroblaster er allestedsnærværende mesenkymale celler med mange viktige funksjoner under utvikling, vevsreparasjon og sykdom. Fibroblaster fra forskjellige anatomiske steder har distinkte og karakteristiske genuttrykksmønstre, men prinsippene som styrer deres molekylære spesialisering er dårlig forstått. Romlig organisering av cellulær differensiering kan oppnås ved unik spesifikasjon av hver celletype; alternativt kan organisering oppstå ved at celler tolker deres posisjon langs et koordinatsystem.
Forfatterne forsøker deretter å komme seg rundt det tilsynelatende paradokset fra romlig spesifisitet i forhold til genomisk ekvivalens ved å stille at 'målsekvenser' - om du foretrekker- molekylære zip-koder - av aminosyrer er ansvarlige for å lede proteiner til bestemte steder i cellen. Men slike 'molekylære zip-koder' skaper ikke et romlig koordinatsystem; de forutsetter det.
Meyer (2004) forklarer det på denne måten:
 I hver ny generasjon oppstår således formen og strukturen til cellen som et resultat av både genprodukter og den eksisterende tredimensjonale strukturen og organisasjonen. Cellulære strukturer er bygd fra proteiner, men proteiner finner veien til å rette lokalt delvis på grunn av eksisterende tredimensjonale mønstre og organisering. innebygd i cellulære strukturer. Preeksisterende tredimensjonal form til stede i den foregående generasjon (enten det er iboende i cellemembran, sentrosomer, cytoskjelettet eller andre funksjoner i det befruktede egget) bidrar til produksjonen av form i den neste generasjonen. Verken strukturelle proteiner alene, eller genene som koder for dem, er tilstrekkelig til å bestemme den tredimensjonale formen og strukturen til enhetene de danner. Genprodukter gir nødvendige, men ikke tilstrekkelige betingelser, for utvikling av tredimensjonal struktur i celler, organer og kroppsplaner (Harold 1995: 2767).
I hver ny generasjon oppstår således formen og strukturen til cellen som et resultat av både genprodukter og den eksisterende tredimensjonale strukturen og organisasjonen. Cellulære strukturer er bygd fra proteiner, men proteiner finner veien til å rette lokalt delvis på grunn av eksisterende tredimensjonale mønstre og organisering. innebygd i cellulære strukturer. Preeksisterende tredimensjonal form til stede i den foregående generasjon (enten det er iboende i cellemembran, sentrosomer, cytoskjelettet eller andre funksjoner i det befruktede egget) bidrar til produksjonen av form i den neste generasjonen. Verken strukturelle proteiner alene, eller genene som koder for dem, er tilstrekkelig til å bestemme den tredimensjonale formen og strukturen til enhetene de danner. Genprodukter gir nødvendige, men ikke tilstrekkelige betingelser, for utvikling av tredimensjonal struktur i celler, organer og kroppsplaner (Harold 1995: 2767).
For et par måneder siden dukket det opp en interessant artikkel i Nature av Zhang et al . For å utløse epitelutvikling ved hjelp av en nål, dokumenterte forskerne at Caemorhabditis elegans, laboratorie-nematode-ormen, er nærliggende celler og deres kontraktile bevegelser uunnværlige for riktig utvikling. Som forfatterne bemerker i sine metode sammendrag:
--For å påføre eksterne krefter på embryoer, ble en nål med en 40 mm stump ende plassert over embryoer som hadde blitt immobilisert på et glassbasert kultur-plate (IWAKI) belagt med poly-lysin og plassert på invertert TCS SP2 konfokalt mikroskop (Leica ). Mikroskopet ble deretter programmert for en tids-forløpssekvens i xyzt-dimensjon med en 6 mm z-avstand, med en 1,6 s-periodisitet for å etterligne pulsen til muskelkontraksjon.
Interessant, i tillegg til å være forankringspunktet for epitelceller, fungerer en struktur kjent som 'hemidesmosomet' også som en 'mekanosensor' som svarer på spenningen indusert av epitelceller nær ved, ved å utløse signalprosesser som til slutt resulterer i celle-type differensiering.
 Vi begynner bare å montere grunnlaget med hensyn til vår forståelse av mekanikken i fosterutvikling og de forskjellige prosessene som ligger til grunn for det. Det er fremdeles mye som ikke er godt forstått om utvikling og et vell av informasjon som fremdeles må læres. Men jeg tror det blir stadig tydeligere at DNA ikke kan inneholde både nødvendig og tilstrekkelig informasjon for morfogenesen av organismeform.
Vi begynner bare å montere grunnlaget med hensyn til vår forståelse av mekanikken i fosterutvikling og de forskjellige prosessene som ligger til grunn for det. Det er fremdeles mye som ikke er godt forstått om utvikling og et vell av informasjon som fremdeles må læres. Men jeg tror det blir stadig tydeligere at DNA ikke kan inneholde både nødvendig og tilstrekkelig informasjon for morfogenesen av organismeform.
Man blir naturlig nok ført til å undre seg over hvordan et så sofistikert system som kontrollerer embryologisk utvikling kunne ha oppstått i kraft av en darwinistisk trinnvis prosess, som det må huskes, tradisjonelt innebærer endringer i DNA-sekvensen. Og vi har sett impotensen av den neo-darwinistiske syntesen på DNA-nivå, ganske atskilt fra lagene med ekstra informasjonsrik kompleksitet som styrer livets show. Altså, for mange moderne darwinister, utgjør ethvert forsøk på å kritisere slike scenarier et 'gud-i-hullene' -argument og bryter det foretrukne prinsippet om metodisk naturalisme.
Selv om oddsen ser forsvinnende tynn ut, blir vi fortalt, og selv om vi for øyeblikket ikke kan tenke oss et gjennomførbart scenario fra Darwin-typen som kunne ha produsert et slikt system, må det likevel være et. Noen evolusjonsbiologer tar tak i dette og tilstår at de er tvunget til å omfavne darwinisme ikke hovedsakelig av vitenskapelige grunner, men av metodologiske (f.eks. Lewontin, 1997). Selv om en slik stilling kan være ganske komfortabel for noen (de trenger ikke noen gang å risikere at deres konseptuelle bygning blir bevist som feilaktig), har jeg rett og slett ikke nok tro til å innta den posisjonen.
Oversettelse og bilder ved Asbjørn E. Lund